染色体的精确分离是保证遗传信息正确传递和基因组稳定的前提,这个过程直接依赖着丝粒区组装的多层动粒蛋白复合体和纺锤体微管间的动态结合。目前,在哺乳动物和酵母中已经鉴定超过100个动粒蛋白,它们之间相互结合形成蛋白亚复合体结构,包括与着丝粒染色质直接结合的内侧组成型CCAN蛋白网络、与微管直接结合的外侧KMN网络复合物、调控细胞周期进程的纺锤体组装检验点(SAC)等。动粒蛋白发生频繁的变异,并与其互作蛋白以多种形式协同进化,导致真核生物动粒组成产生复杂的多样性。因此,在植物中对动粒蛋白组成及其生物学功能进行研究有助于阐明真核生物动粒复合体的复杂进化历程,对理解染色体正确分离和取向的基础生物学过程提供不同的认识。
韩方普研究组科研人员从玉米 (Zea mays) 中克隆了 Knl1 同源物,并将其确认为组成性中心动粒成分。功能测定证明了它们在核分裂过程中的染色体聚集和分离中的保守作用,从而导致当 Knl1 转录物耗尽时内核发育过程中的细胞分裂缺陷。玉米 Knl1 中间的一个 145 aa 区域,不涉及 MELT 重复,与纺锤体组装检查点 (SAC) 组件 Bub1/Mad3 家族蛋白 1 和 2 (Bmf1/2) 的相互作用有关,但与 Bmf3 无关蛋白质。它们可能与 Bmf1/2 的 TPR 结构域形成具有疏水界面的螺旋构象,类似于脊椎动物的结构。然而,在单子叶植物中检测到的这个区域在双子叶植物中表现出广泛的差异,表明 SAC 与动粒连接的不同模式存在于植物谱系中。这些发现阐明了 KMN 网络在细胞分裂中的保守作用以及 在SAC 信号和动粒网络中进化模式的显着动态。本结果为动粒蛋白网络和SAC信号通路之间提供了一个协同进化的模式。
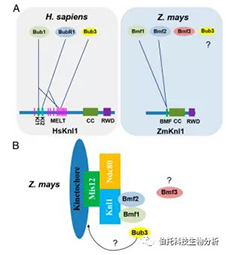
人类和植物中具有 SAC 信号的 Knl1 结构模型。
(A) Knl1 蛋白的结构域与人类 (H. sapiens) 和玉米 (Z. mays) 之间的 SAC 信号结构的比较。在人类中,Knl1 MELT 重复与 KI 基序合作以募集 SAC 信号(Bub1, BubR1 和 Bub3)。在玉米中,Knl1 的 MELT 重复序列丢失,Knl1 中间的 145 aa 区域参与了与玉米中 Bmf1 和 Bmf2 的相互作用。然而,在 Knl1 和 Bmf3 或 Bub3 之间没有检测到相互作用。Knl1 上不同颜色的矩形代表 MELT、KI1、KI2、BMF(BMF 结合域)、coiled-coil(CC) 和 RWD 域。实线代表检测到的与 Y2H 系统的交互。(B) 在玉米中检测到 Bmf1 和 Bub3 的细胞周期依赖性动粒定位,这是 SAC 信号传导的典型特征。Bmf1 和 Bmf2 的 TPR 域是它们与 Knl1 相互作用所必需的。Bub3 如何被招募到动粒以及 Bmf3 在玉米中的作用仍然未知。


(C) ZmKnl1-M8 和 ZmBmf1/2/3 之间相互作用的荧光素酶互补成像测定。荧光信号强度代表它们的相互作用活动。
*:本文献中使用了德国伯托的植物活体影像系统Berthold NightOWL。
文章链接:
https://doi.org/10.1073/pnas.2022357118
